網膜と第一次視覚野 V1 の間には連続的な 1 対 1 対応が存在します retinotopy。 鼓膜の周波数選択特性と第一次聴覚野 A1 との間にも対応関係が見られます tonotopy。 同様に体表面の感覚と体制感覚野の間にも対応関係が見られます somatopy。 すなわち感覚器官と第一次感覚野との間の神経結合は、 類似した刺激に対して皮質上の同じような位置に対応する受容野を持つことが知られています。このような 2 つの神経場間の連続的な結合関係のことを トポグラフィックマッピング topographic mapping と言います。
視覚野のトポグラフィックマッピングについては、さらに細かいことが分ってい て任意の視覚位置に対して、眼優位性 ocular dominancy, 方位選択性 orientation selectivity, 色 などの情報が処理されるように規則正しく配列さ れています。ハイパーコラム hypercolumn 構造といいます。 ハイパーコラムは、2 次元しかない皮質上に、2次元の網膜位置、 方位、視差情報(立体視)、色情報処理などの多次元情報をなるべく効率よく 処理しようとする生体情報処理の機構を表していると言えます。
このような構造は、
大まかな構造は遺伝子によって決定されますが、
細かい構造については神経回路の
自己組織化 self organization によって達成されると考えられています。
このような例は枚挙に暇がありません 。
一例として Linsker の行なったシミュレーションを紹介します。
。
一例として Linsker の行なったシミュレーションを紹介します。
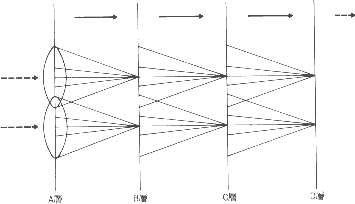
Linsker は図2 のような数層の細胞で構成されるモデルを考え ました。
各細胞層では、多数の細胞が 2 次元的に広がっています。 B 層の各細胞は A 層の細胞から入力を受けます。たとえば、 A 層の円で囲まれた領域にある細胞 100 個がそれぞれ B 層の 一個の細胞に結合しています。同様にして、B 層から C 層、C 層から D 層 へと続く結合も同様です。結合強度の変化は Hebb の学習則の変形(表 1)で 入力と出力ともに活動度が高い場合には結合を強める他に、 両方ともの活動が低ければ結合の強さを弱めるような学習です。
入力は A 層から入りますが、A 層においてはランダムで相関がない活動が自発 的に起きていると想定しました。その結果、次のようなことが起きました。A か ら B への結合では、結合の強さは正の大きな値になるか、負の大きな値になる かのどちらかでした。結合が正の興奮性になった場合にどういうことが起きたの でしょうか? そのとき細胞が観察しているのは、細胞層 A のある領域の入力の 平均です。B 層では、隣り合う細胞が受け取る A 層の細胞の範囲は重なりあっ ています。したがって、この二つの細胞は A 層におけるパターンの似た部分を 見ており、ある時刻に一方の細胞の活動度が高ければ、もう一方の細胞の活動度 も高い可能性がたかいです。つまり、B 層において、近くに位置する 2 つの細 胞の活動度の間には相関が生じています。
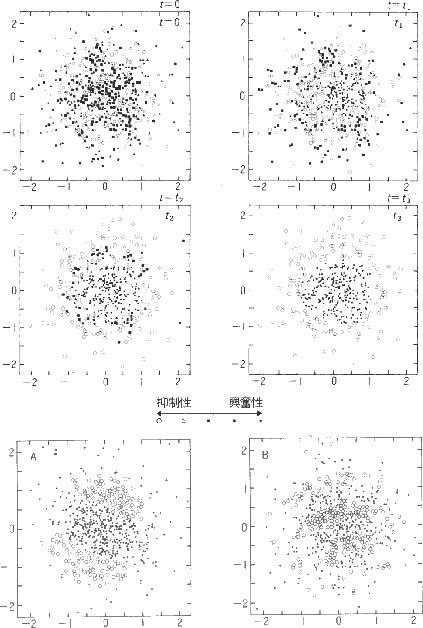
次に B 層から C 層への結合の変化が図3 に示されています。
図3 では C 層の一つの細胞を取り上げ、そこからB 層 をながめています。つまり C 層の一つの細胞へ入力する B 層の細胞を見ている わけです。シミュレーション開始時(t=0)にはすべての結合はランダムですが、 時間が経過するにつれて中央に点が見えてきます(興奮性の領域)。周辺には 抑制性の領域が現れます。 ランダムな結合と Hebb 則だけを使って、 on 中心型細胞が生じることが 示されたことになります。さらに先の層には方位選択性の細胞が現れます。
Linsker の提案したネットワークを追試するのは、非常に簡単なプログラムで可
能です。にもかかわらず、第一次視覚野で
の細胞の特徴をよくとらえた優れたデモンストレーションと言えます。
自己組織化の特徴を抽象化してとらえれば、多次元刺激をその刺激の持つ規則性に 従って 2 次元の皮質上への対応問題ととらえることができるます。 入力層の空間多次元多様体から 2 次元部分空間への写像といいます。 このような立場から、統計学の用語を用いて説明を試みます。